Instructor: Dean Gabriel
It is useful to understand the concepts that developed over time to explain heredity in populations; many were developed prior to Mendel's publication of the concept of genes as a unit of heredity, and certainly prior to the idea that mutations in genes arose spontaneously. Biologists began to wrestle with the concept of how populations of a given species were maintained, once biologists begin to appreciate that species could change over time. Prior to that, species were thought of as immutable (unchanging) acts of final creation and maintained in a more or less static way over time. After all, the world was only a few thousand years old, (Bishop Usher had declared that God created the world in 4004 BC), and there was no need for anything to have changed since Adam named the species...
Yet breeders of animals and plants had changed the appearances of, for example, dogs and crop plants and the cataloguing of species was proving more difficult than imagined. Did species change or evolve? How? What sort of variation is observed within a species? How much variation can be tolerated before two species are recognized, or until a new species evolves from the old?
The first scientifically testable theory for the origination of variation within a species is now generally credited to Jean-Baptiste Lamarck. In 1809, he published the theory that the environment caused organisms to acquire characteristics that were heritable. His "First Law" was that use or disuse causes structures to enlarge or shrink ("what you don't use, you lose"). For example, people living at high altitudes and Japanese pearl divers have an expanded lung capacity (larger lungs); the eyes of organisms raised completely in the dark tend to atrophy; immunity against severe diseases can be acquired by vaccination, etc. His "Second law" stated that all such changes were heritable. Therefore natural selection itself caused changes in a population. This theory is best summed up as the theory of inheritance of acquired characteristics. In modern day terms, a Lamarkian form of inheritance would be the reverse of DNA makes RNA makes proteins makes function. As with all good scientific theories, this theory was testable (eg., do the offspring of Japanese pearl divers have larger lungs if they don't dive?).
By contrast, Charles Darwin in 1859 stated that natural selection of the most fit pre-existing variants caused changes in a population. Again, neither Darwin nor Lamark knew about the concept of "genes", let alone the concept of genes residing on chromosomes, let alone that genes are encoded by DNA. Both Lamarck's and Darwin's views seemed cold, or even cruel, and certainly directionless. They appeared to violate religious beliefs, and they were not well accepted. Both men drew public ridicule, and Lamarck died destitute. But the scientific theories of both were testable.
Gregor Mendel published his work defining genes as a unit of inheritance and the laws of independent segregation and assortment in 1865, but it was not until the year1900 (after his death) that his publication became widely known. Hugo de Vries and others independently discovered Mendel's previously obscure work, published in a local Austrian naturalist society, and advertised it widely. de Vries went further and proposed that mutations alone were the major source of evolution and new species. To him, the environment acted upon variation in individuals, but did not affect the genes. Therefore characteristics could neither be "acquired" nor "selected" from those which are pre-existing, they simply occurred. This idea was called "mutationism" or "Mendelism", and it was embraced by many, especially those thinking at the organismal or cellular level. After all, the genes became known to reside on chromosomes and mutations were induced in genes, and therefore, random favorable mutations could be the major source of gene change. The evidence that mutations can occur independently of environmental influences and as a matter of chance was demonstrated adequately in Drosophila (and other organisms). The demonstration by H.J. Muller that lethality was more commonly induced than any other type of mutation simply meant that random favorable mutations were much more rare than random unfavorable ones. Proof that selection was based on genotype was provided at the population level by mathematicians and biometricians such as R.A. Fisher, J.B.S. Haldane and Sewall Wright in the 1920s and 1930s. The fact that selection was based on genotype did not settle the question of how selection operated.
A major contribution to the debate between the Lamarkianists and Darwinists was made by August Weismann, who provided evidence and argued vigorously that the germ cell line was independent of somatic cells in higher organisms. This conceptual breakthrough was critical in the debate against Lamarckianism, since it could explain, if true, why body changes produced in a parent by the environment were not inherited in the offspring. "The germ-plasm of a species is never generated de novo; it only grows and multiplies continually, handed down from generation to generation..." "The sought-for bearer of heredity is contained in the chromosome material" (A. Weismann, 1904, Vortrage uber Deszendenztheorie, Jena 198). Weisman argued that the parent's somatic cells, which are changed by the environment, were not the source of the hereditary material, and that the hereditary material in the chromosomes was not ordinarily affected by the environment. Proof for these assertions was more difficult, however.
Weisman tried to disprove Lamarckian theorists in various ways, but provided no serious tests of the theory. One of Weisman's experiments involved cutting off the tails of mice, generation after generation, but this proved nothing, since there was no environmental need for losing the tail. (Perhaps he realized that Hebrews had been circumcising male infants for thousands of years, but Jewish boys were still born with foreskins). Weismann called his theory "Neo-Darwinism", and it is widely accepted today (although the proof was by others). Neo-Darwinism or the "synthetic theory" is that mutations occur randomly in the germplasm, and natural selection of the individual that develops from the germ line determines whether the mutations are favorable or not, by virtue of how they affect the reproductive rate of the individual that carries that germplasm. This theory preserves the essential Darwinian concept of natural selection, and explains why somatic cell alterations in a parent do not affect the offspring of the parent.
One of the earliest critical tests of Neo-Darwinism vs. Lamarkianism was by Luria & Delbruck. (Click on the link)
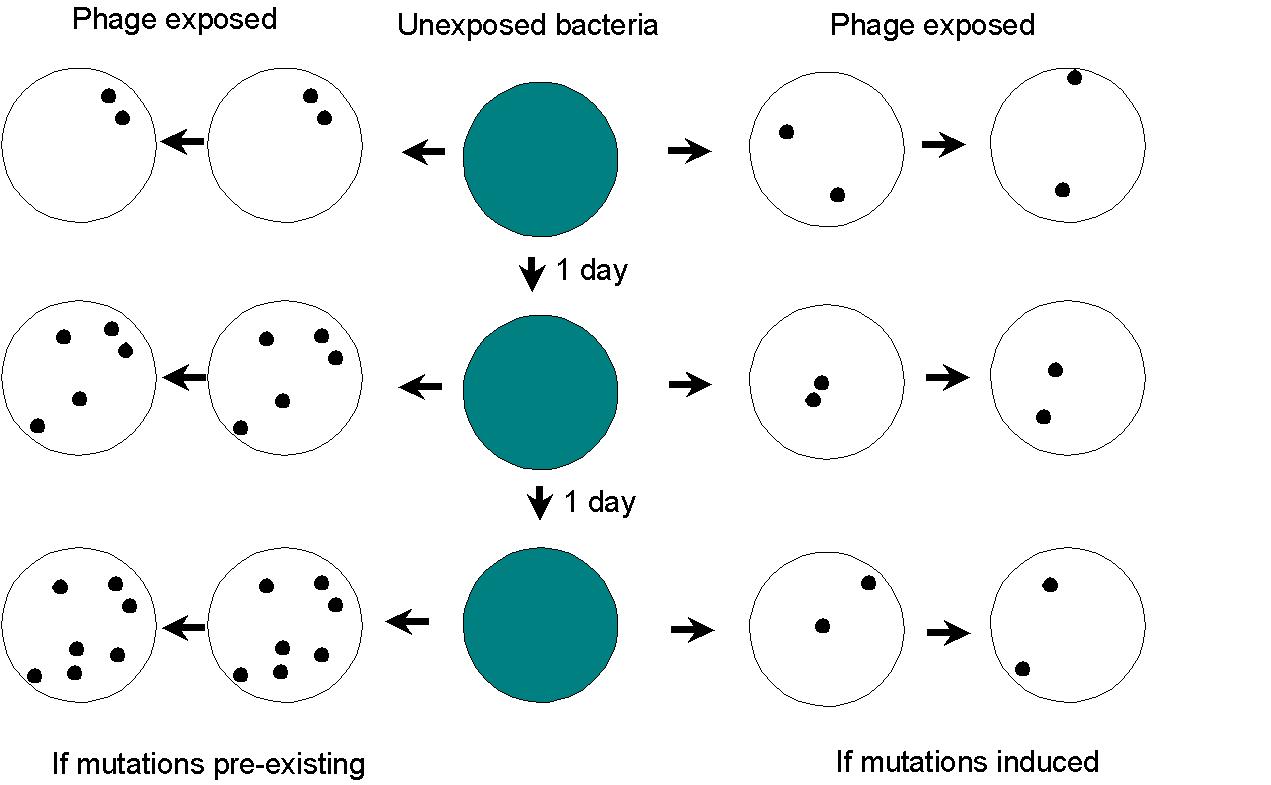
A simple method of proving that mutations were pre-existing or pre-adaptive,
as opposed to being induced by the environment,
was provided by Joshua Lederberg using replica plating: Basically,
a lawn of bacteria is replicated over a period of several days. If
mutation rates are constant and pre-existing, the earliest plates will
have fewer mutations, but they will stay localized as they are replicated.
Even duplicated replica plates will show some bacteria, mutant in the original
plate, in the same location. However, if mutations are induced by
exposure, then each mutation should occur in a different location, and
not accumulate over time.
The conclusion was, again, that mutations occur independently of selection.
H.J. Muller had proven that when he induced mutations with X-rays, by far the
most common mutations were lethal. The second most common class of mutations were those with some
discernable deleterious effect---they caused abnormalities that reduced the chances that individuals
carrying the mutations would live to reproductive age, and even if they did live to reproductive age,
would be less likely to reproduce, and if they did reproduce, would be less capable and therefore less
likely to assist in the survival of their own offpsring (in cases where such assistance is needed),
For example, if a dominant abnormal allele leads to early death, such that an individual with the allele
produces on average 1/4 of the offspring of a normal individual, then the frequency of the abnormal allele
in the population should decrease by 75% with each generation, then in only four generations, the frequency of such an allele will be less than 4/10 of a percent of the population. Why do we then see a relatively steady rate of births in all populations of particular abnormal alleles?
The answer is recurrent mutations. The rate of such mutations can be estimated, if we assume an equilibrium between the number lost due to reduced fitness and the number gained due to mutation. Let N be the total number of individuals of one generation, x the frequency of the abnormal allele among them, and w the relative reproductive fitness of the allele. Let r be the mutation rate or frequency per gamete of mutation from normal to abnormal.
Total parents in the population: N
Assuming a diploid population and autosomal (not sex linked) gene locus, there are xN affected parents who have xN abnormal and xN normal alleles.
There are (N - xN) normal parents who have 2(N - xN) normal alleles.
The sum of all normal alleles is: xN (from affected parents) + 2N - 2xN = N(2-x).
If mutations from normal to abnormal occur at rate r, then number of new mutations is: rN(2-x)
If the frequency of the abnormality is very low (say, 1/10,000 or smaller) in the population, we can neglect x and simplify the number of new mutations to: 2rN
The number of abnormal individuals lost in each generation is the fraction lost due to relative reproductive fitness, or 1 - w (1 - w is also called the coefficient of selection, s), times the number of abnormal alleles present, xN
The number of lost alleles is therefore: (1 - w)xN
For equilibrium between lost and gained mutant alleles, 2rN = (1 - w)xN
Solved for mutation rate, r = 1/2(1 - w)x
Reproductive fitness and frequency of an abnormality are measureable, giving an estimation of mutation rate.
The following example is from Curt Stern's Principles of Genetics:
Achondroplastic dwarfism was studied in Denmark. A total of 108 dwarfs produced 27 children. Over the same time period, 457 non-dwarf siblings of the dwarfs produced 582 children.
Calculate the relative reproductive fitness w and selection coefficient s of the dwarfism allele.
There were 10 dwarfs born in the population among 94,075 live births. Calculate x, the frequency of the abnormality. What is the mutation rate for dwarfism?